|
DEVELOPMENTAL BIOLOGY 3230 |
|
|
|
History and Basic Concepts, Principles of Development pg. 1-29 |
|
|
|
 |
|
|
|
 |
Aristotle suggested that embryos were either preformed in the "embryo" and just grew in size, or they formed from something simpler via epigenesis. For religious reasons many wanted to beleive that all embryos were created with and contained in the first "man". The earliest microscopists drew pictures such as the one at left suggesting that they could see the preformed embryo in the head of the sperm. This preformation idea was laid to rest with the advent of better microscopes and the growing evidence that all animals are made up of cells. The Cell Theory (1820-1880) advanced by Schleiden and Schwann proposed that all animals were communities of diversely functioning cells. The egg was identified as a single large cell and observed to divide during embryogenesis. |
|
|
|
 |
|
|
|
 |
|
|
August Weismann in the late 1800s put forth the theory of germ plasm. The important advance was the distinction between somatic cells and germ cells. Inheritance is solely through "germ" cells: the gametes (sperm and egg). Futhermore, this period saw the advance of Darwin's idea of evolution through natural selection combined with the re-discovered experiments and ideas of Gregor Mendel. Microscopists observed chomosomes in cells and described how there was an equal contribution of male (sperm) and female (egg) chromosomes to the zygote. This combined chromosome set was then seen to replicate and pass on to all of the cells of the embryo. |
|
|
|
 |
|
|
Charles Darwin took his trip on the Beagle when he was 22 years old!! |
|
| The figure above illustrates the distinction between germ cells and somatic cells. Mutations (or any environmentally induced changes) in the somatic cell do not get transmitted to the next generation. Inheritance is strictly through the germ cell lineage.
The figure below illustrates another idea proposed by Weismann, that nuclear "determinants" are unevenly segregated to different cells during the cleavage stage of development and specific different cell fates. This was consistent with the results of experiments performed in tunicates and other animals where development is "mosaic".
|
|
|
|
 |
|
|
 |
|
|
 |
|
 |
Matthias Schleiden and Theodor Schwann, founders of modern cell theory. The cell is the basic unit of life. All animals are made up of communities of cells. |
|
|
|
Gregor Mendel |
|
|
|
 |
|
|
 |
|
|
Wilhelm Roux perform this "ablation" experiment to test Weismann's therory of "determinants" and mosaic development. The results of ablating one cell of the two cell embryo was a half embryo. This was consistent with Weismann's prediction of localized determinants and mosaic development. The result was correct, but as we shall see the interpretation was not. If you do the "same" experiment by separating the two blastomeres you get a very different result---- two perfectly formed smaller tadpols!! |
|
|
 |
|
|
| Hans Driesch tried a similar experiment in the sea urchin, but got dramatically different results. If he seprated the two blastomeres he got perfectly normal development of half sized larva. This result was NOT supportive of the mosaic model of cell fate specification. The half embryo (1 cell of a two cell embryo) was able to "regulate" and develop without any missing parts. This was the first compelling example of the important developmental process of REGULATION. Think about how Driesch's and Roux's experiments differed in design if not in intent. How would you explain the different results? What is more compelling a positive result or a negative result? |
|
|
|
 |
|
|
|
 |
|
|
| The importance of cell-cell communication in development was dramatically demonstrated by a transplantation experiment done by Hans Spemann and Hilda Mangold (grad student!). They demonstrated that transplanting one particular region of a donor embryo, the dorsal lip of the blastopore, to an ectopic location in a host embryo they could "induce" a new secondary embryo. They demonstrated that the new embryo was made up mostly of host cells, not the donor cells! This meant that the donor dorsal lip cells somehow "induced" the nearby host cells to change they fate and take part in the formation of a duplicate embryo. This dramatically demonstrated that cell-cell communication is important for regulating cell fate and that one small group of cells can act as an "organizer" of embryonic pattern formation. Hilde Mangold die tragically in an accident (home furnace malfunction) before she could be honor for her achievement. Spemann received the Noble prize for this discovery. |
|
 |
|
|
|
 |
|
|
 |
|
|
 |
|
|
|
Wilhelm Johannsen (seated left in garden, Bateson seated right) in 1909 made the important distinction between genotype and phenotype. It seems obvious now that the genotype is the gene compliment inherited from you parents, while the phenotype is the observed structure, physiology, behavior, etc that an organism exhibits. It is clear from this picture of identical twins (same genotype) that the phenotype can be influenced by epigenetic and environmental factors. |
|
|
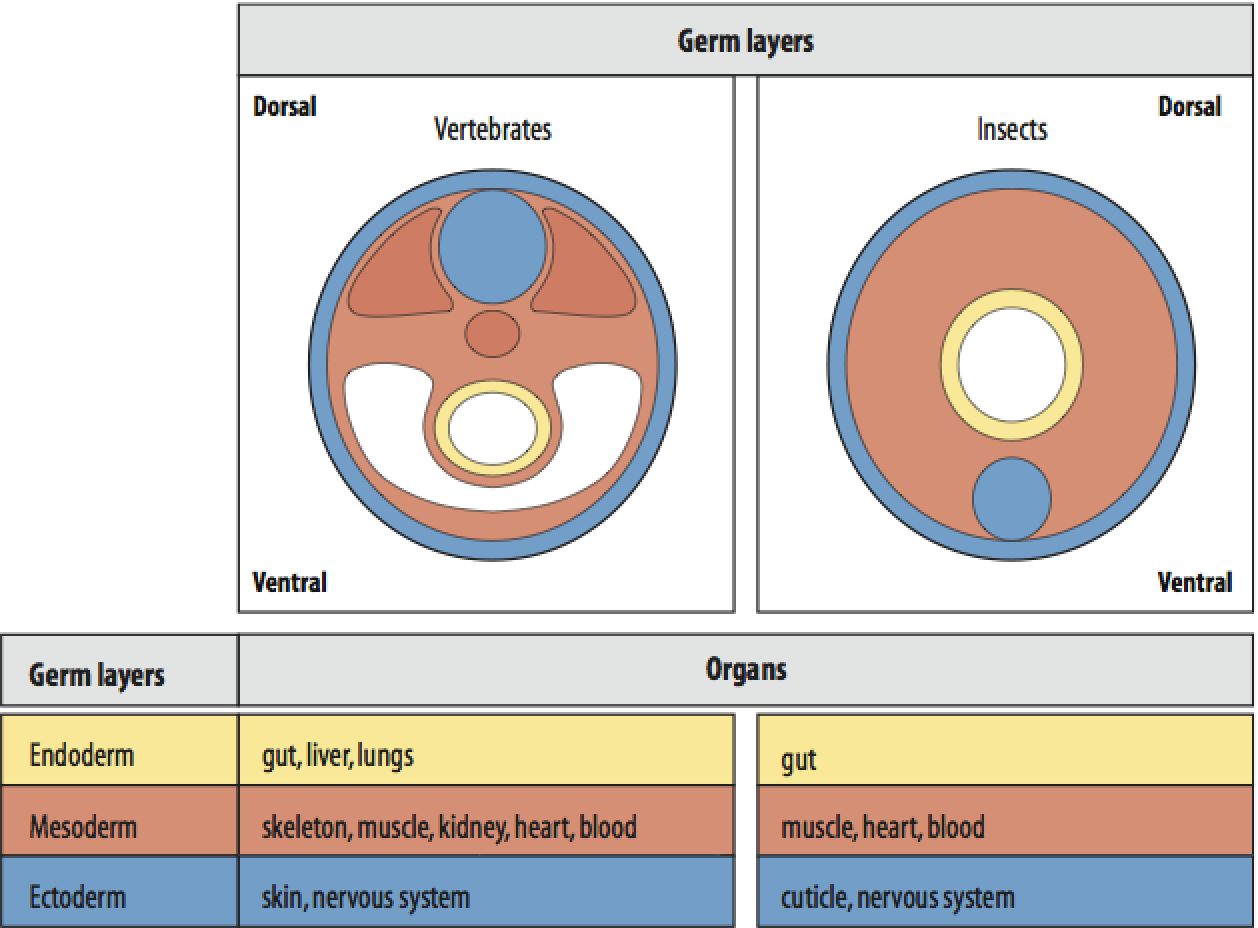
Pattern formation is a key concept that is intensively studied by developmental biologys. The earliest and most important patterns that emerge during embryogenesis are the whole embryo axial patterns shown below. Anterior-Posterior, Dorsal-Ventral, and Left-Right. The axial patterning visible on the surface of the embryo is occuring concurent to the patterning of the internal tissures shown at right. The cell movements during gastrulation not only make the axial patterns visible, but are also important for the formation of the three "germ" layers, outer ectoderm, middle mesoderm, and inner endoderm. |
|
|
 |
 |
|
|
Morphogenesis, or a change in form, is first and most dramatically seen during gastrulation where the hollow ball of cells (one layer) is transformed into a three layered embryo (ectoderm, mesoderm, and endoderm. In the picture below after gastrulation (far right) all the cells on the surface are ectodermal, the inner tube layer of cells opening to the mouth and anus are endoderm. Cells that migrate between the inner and outer tubes become the middle mesoderm layer (not shown). |
|
|
|
 |
|
|
 |
|
|
Individual cells can communicate and act either as a co-operative unit or as individuals. To the left is shown an example of cells in an epithelial sheet that are joined by cell-cell junctions that provide both physical and "informational' connections. Individual cells can change the arrangement of their cytoskeletal proteins and cause the whole sheet of cells to bend as a unit. Thus the behavior of individual cells working together can lead to a macroscopic change in form. Cells can migrate as individuals or in sheets. During development there are many instances of epithelial to mesenchymal transitions where cells in an epithelium egress and migrate as individual and then reassociate to form organs, eg., heart formation. |
|
|
|
 |
|
|
|
Embryos generally grow in cell number and size during development. Notice, however, that there is often large changes in proportion during development. Notice in the human embryo that the head develops early and is relatively larger at earlier stages. Human development occurs primarily in utero where the embryo does not have to "function". Thus development only has to ensure the most favorable end product and can build structures in the order it wants without needing the embryo to be functional from the earliest times. |
|
|
|
Conceptually, there are three main types of experiments that developmental biologist do to understand the timing and mechanism of cell fate specification and determination. They are cell ablation, cell transplantation, and cell isolation. Normal cell fate is simply the fate of a cell at a particular position in a normal undisturbed embryo and relies on the stereotyped cell movements that occurs during embryogenesis. An embryo cell fate map tells you nothing about how or when cell fate is specified or determined. When Region B is transplanted to a new location and takes on the fate of cells at that new location we say that AT THIS TIME Region B's fate has not been determined. In the next transplant we see that when Region B is moved to a new location it now differentiates based on its original location. We say that AT THIS TIME Region B's fate has been determined. In the last example we isolate the cells of Region B and they differentiate based on the location they were taken from in the embryo. We say that AT THIS TIME Region B has been specified. What ablation experiments could you do to learn about when and how cell fate is determined? |
|
|
 |
|
|
|
 |
|
|
Shown here is a real example using transplatation to define the eye field and when it is determined. Note the use of a donor and host. The developmental stage of both the donor and host can be varied, in addition to the positions of the graft. This allows one to identify the source and timing of potential inductive signals. |
|
|
 |
|
|
|
Please review the regulation of gene expression and protein synthesis. You should be familiar with the terms in the figure and have a basic understanding of the regulation that can occur at each numbered step. At one level, all of development is just understanding the pattern of gene expression in all cells of the embryo throughout development. This is obviously a gross simplification since proteins are functional machines and signaliing molecules that can interact in complex ways. However, we will see that much early axial patterning is first indicated by stereotyped patterns of expression of transcription factors. Transcription factors are proteins that bind directly to DNA enhancer/promtor regions (or to other transcription factors that bind to DNA) and regulate gene expression. They can act as molecular switches setting the pattern of expression of many genes. |
|
|
Here are some simple examples of gene regulatory "circuits". In the first example of Positive Feedback you see that the activator (a transcription factor) binds to the enhancer/promotor region of gene 1 and activates transcription. The protein product of gene1 (another transciption factor) binds to the enhancer/promoter region of gene 2 and activates transcription. Now the protein product of gene 2 (transciption factor) feeds back by binding to and activating transcription from gene 1. Thus a single transient signal from the activator leads to a permanent "on" state for gene 1 and 2. You no longer need the activator signal.
In the second example of Negative Feedback you can see that the activator signal turns on gene 3, which activates gene 4. The product of gene 4 feeds back onto gene 3 and inhibits its transcription. Thus a single transient signal from the activator is turned into a transient signal from this gene circuit.
Now you can imagine how complex patterns of gene expression can permanently be effected in cells by single "activators".
|
 |
|
|
|
As we discussed earlier, Induction is the process by which one cell or group of cells influences the developmental fate of another group. Inductive signals are defined experimentally as either Permissive or Instructive (What experiments would you do to distinguish a permissive from an Instructive signal?). Inductive signals can be transmitted to cells via Diffusion, Direct Contact between a cell surface ligand and receptor, or it can diffuse directly between cells via Gap junctions. Secreted molecules can diffuse over long distances to provide more "global" inductive signals. It is also possible for the same signal to have different effects depending on "threshold" concentrations and the identity of the receptor expressed by the target cells. Moreover, the same signal can have different effects on the same cell at different developmental times (How?). |
|
 |
|
|
|
 |
|
 |
|
|
|
The French Flag Model of pattern formation illustrates an important concept in developmental biology. A group of cells that has the same "potential" (an equivalence group) is exposed to a gradient of a morphogen. (A morphogen is generally a secreted and diffusable inductive signal). Thus the cells closest to the source of the morphogen are exposed to the highest concentration and those farthest away to the lowest concentration. The cells of the equivalence group are see a continuously varying concentration of morphogen depending on their POSITION in the "embyronic field". If we imagine that there are threshold values for different inductive signals then we can see how this single morphogen gradient can impose a "digital" pattern of cell fates on the cells of the equivalence group that make up an embryonic field. (What characteristics of the morphogen and it cognate receptor(s) might lead to these threshold effects?) |
|
|
The concept of positional information is very powerful. It can be used to generate very complex patterns of cell behavior. Just as positional information can be used to specify almost any word by this crowd in the stadium, so you can imagine that using positional information to specify which genes are expressed by cells in an embryonic field can generate complex patterns of morphogenesis and function. |
|
 |
|
|
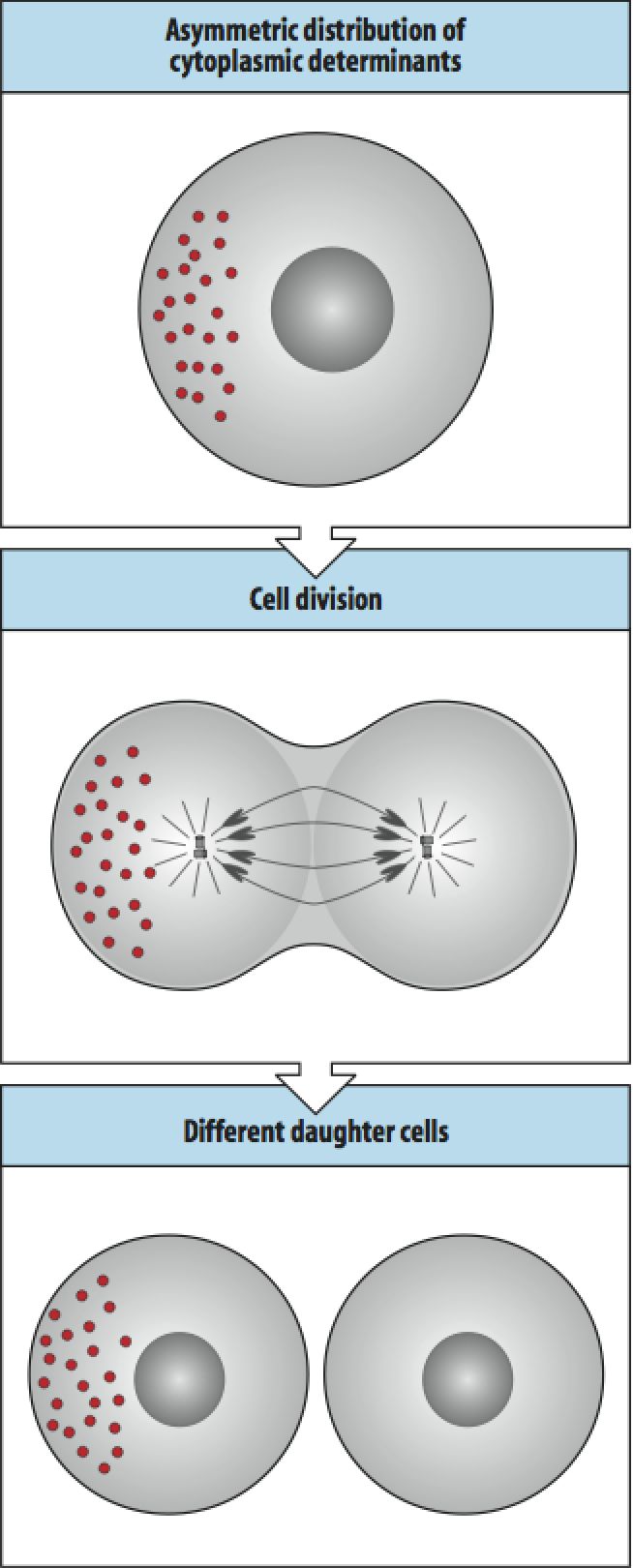
| The asymmetric distribution of cytoplasmic determinants is similar to Weismann's idea, but distinct in saying that it is NOT the DNA that is differentially distributed, but cytoplasmic informational molecules (protein, mRNAs, etc.). Note that in the example, one of the daughter cells is just like the "mother" cell and one daughter is different. Thus these two cells could have very different potentials for differentiation. This pattern of cell division with the asymmetric distribution of a localized cytoplasmic determinant is thought to occur in stem cell lineages as described below. The "stem" cell division produces a new cell fate and regenerates the orginally "stem" cell fate. |
|
 |
|
|
Stem cells are defined as cells that are pluripotent, that is they can give rise to cells with several possible fates. Stem cells can undergo several patterns of cell division as shown at right. The stem cell can undergo several rounds of mitosis where it generates a novel daughter cell and regenerates itself (S to X). It can divide and generate more stem cells (S to S) or it can divide and produce two daughters that can take on two different fates different from the stem cell (S to X and Y). |
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|